Варианти на изпълнения Toll-подобни рецептори. NOD-протеини
TLR5 (1q33), експресиран на клетки от далака и периферни кръвни левкоцити, ендотелиални клетки. TLR5 признава флагелин Грам-положителни и Грам-отрицателни бактерии и спомага за индуциране на сигнална каскада, включваща хемокини, цитокини и костимулаторни молекули.
TLR6 (4р) експресира върху периферни кръвни левкоцити и клетки от далак образува димери с TLR2 и признава, пептидогликан и зимосан.
TLR7 (Hr22.3) се експресира в клетки на белия дроб, плацентата, далака и периферни кръвни левкоцити. Естествената лиганд за TLR7 все още не е намерен. Работа извършва на TLR7-дефицитни мишки са показали, че TLR7 признава имидазолови съединения, като R848, които имат антивирусни свойства, и системно прилагане на TLR7 агонист осигурява антивирусен ефект.
TLR8 (Hr22) идентифицира с TLR7 и TLR9, изразена в големи количества в периферни кръвни левкоцити и клетки на белия дроб.
За да се изследва функционалната роля TLR7 и TLR8 и активируем репертоар на вродени имунни клетки, групата КА Гордън използват селективни агонисти на тези рецептори. TLR7 агонисти пряко активират плазмацитоидни DCs пречистени, и в по-малка степен моноцити. Обратно, TLR8-агонисти повишени активирането на DCs миелоидни и моноцити. Съответно, TLR7-агонисти са по-ефективни в сравнение с TLR8-агонист за предизвикване на IFN-а и IFN-регулиране цитокини (IFN-индуцируем протеин и IFN-индуциран Т-клетъчен хемоатрактант MLPK човек). Агонисти на TLR8-индуцирани провъзпалителни цитокини и хемокини (TNF-а, IL-12, и M1R-1А).
Въпреки това, естествен лиганд за TLR8 Все още не е открит, подобно чувствителни към TLR7 имидазолови съединения с антивирусна активност. Също така разкрива, че TLR8 играе ключова роля в разпознаването на едноверижни РНК вируси (Coxsackie В и parechovirusa 1).

TLR9 (Zr21.3) се локализира вътреклетъчно в ендоплазмения ретикулум, последвано от изместване след стимулиране в сомата ендо TLR9 участва в разпознаването на неметилиран CpG-мотиви на бактериална ДНК, което е доказано в експерименти с мишки и клетъчни линии с дефицит на TLR9. Те анулиране всички CpG-индуцирани ефекти (производство на цитокини, костимулаторен молекули-ING, лимфоцитна пролиферация).
TLR10-23 (4r14) отвори наскоро. Въпреки това, техните функции и лиганди все още остават неопределени. Е установено, че те са преобладаващо експресиран в клетки на имунната система в по-голяма степен на В-лимфоцити и DC plazmotsitoidnyh. Смята се, че TLR10 и други Toll-подобни рецептори, участващи в имунния отговор, и могат да действат като съ-рецептор, като TLR1 и TLR6.
Toll-подобни рецептори признае консервативни молекулни продукти от различни класове на патогени, включително грам-положителни и грам-отрицателни бактерии, ДНК и РНК вируси, гъбички и протозои. Разпознаване на TLRs лиганди води до развитието на поредица от сигнали, които предизвикват бързото имунен отговор, необходим за убиване на патогена. Сигналите, получени от TLRs, също са отговорни за индукция на DC съзряване и последващо индуциране на адаптивен имунен отговор.
NOD-протеини. Последните изследвания са показали наличието на други вродени имунни рецептори за разпознаване, т.нар нуклеотид-свързващ домен oligomerizing (нуклеотид-свързващ домейн олигомеризация - NOD) протеини, които функционират в цитоплазмата на клетки-гостоприемници. NOD рецептори са включени в признаването на бактериална пептидогликани вътреклетъчен [263, 640].
NOD2 признава запазен muramilpeptidnye клетъчна стена структура на всички бактерии, докато NOD1 признава диаминопимелинова киселина присъства в пептидогликановия клетъчната стена на грам-отрицателни бактерии. Както TLRs, функционално значение NOD-сигнал се идентифицира при пациенти с NOD-мутации, които са по-податливи на хронични заболявания на бактериален произход.
 Алцхаймер научих за откриване на луминесценция на левкоцити
Алцхаймер научих за откриване на луминесценция на левкоцити Свойствата и характеристиките на Т-супресори. Взаимодействието на Т-супресори
Свойствата и характеристиките на Т-супресори. Взаимодействието на Т-супресори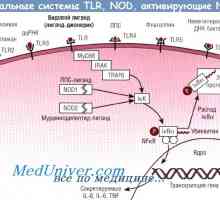 Изпълнения obrazraspoznayuschih PRRS рецептори. Toll-подобни рецептори
Изпълнения obrazraspoznayuschih PRRS рецептори. Toll-подобни рецептори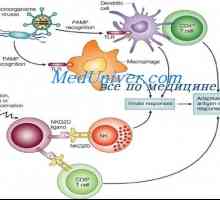 CpG неметилирана ДНК мотиви. Lipoarabinomannan, РНК двойна спирала,…
CpG неметилирана ДНК мотиви. Lipoarabinomannan, РНК двойна спирала,…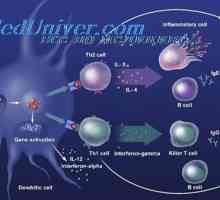 Протеини на топлинния шок (HSPs). Ефектите на вродения имунен отговор към HSP,
Протеини на топлинния шок (HSPs). Ефектите на вродения имунен отговор към HSP,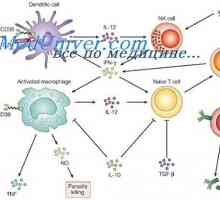 Лиганди на рецептора на ефектори на вродения имунитет. Пептидогликан липопептидите
Лиганди на рецептора на ефектори на вродения имунитет. Пептидогликан липопептидите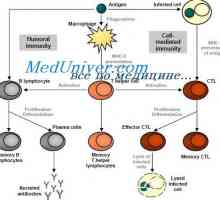 Сигнали дан-подобни рецептори. Ролята на молекула MyD88
Сигнали дан-подобни рецептори. Ролята на молекула MyD88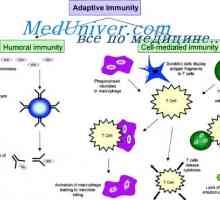 Лимфоидна или плазмацитоидни дендритни клетки. Функцията на дендритни клетки
Лимфоидна или плазмацитоидни дендритни клетки. Функцията на дендритни клетки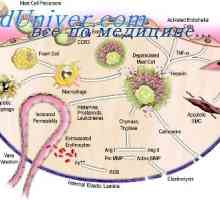 Взаимодействието на Т-хелперните клетки с дендритни клетки. Диференциацията на CD4 клетки
Взаимодействието на Т-хелперните клетки с дендритни клетки. Диференциацията на CD4 клетки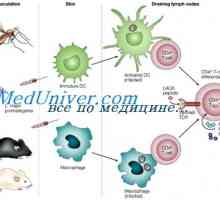 Интерлевкин-13 (IL-13). Трансформиращ растежен фактор (TGF)
Интерлевкин-13 (IL-13). Трансформиращ растежен фактор (TGF)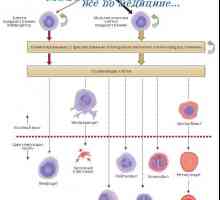 Видове бели кръвни клетки. Произходът на белите кръвни клетки
Видове бели кръвни клетки. Произходът на белите кръвни клетки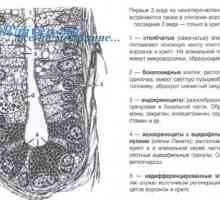 Епителиалните контролни бактерии в червата: Мамре и PRR
Епителиалните контролни бактерии в червата: Мамре и PRR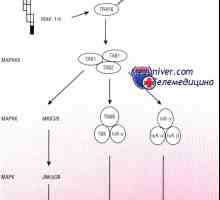 Имунният отговор на епител и бактерии активиране път на апоптоза чревната
Имунният отговор на епител и бактерии активиране път на апоптоза чревната- Взаимодействие с чревна микрофлора и тяхното взаимно влияние
- Ниските нива на левкоцити причини левкопения
 Съставът на CSF
Съставът на CSF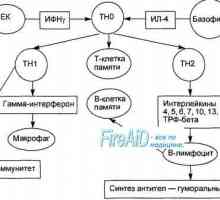 Активирането на Т и В лимфоцити в имунния отговор. Активирането на лимфоцити. Образува специфичен…
Активирането на Т и В лимфоцити в имунния отговор. Активирането на лимфоцити. Образува специфичен…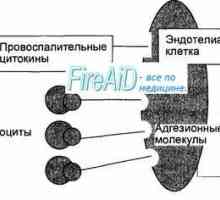 Етап имунен отговор. Форми на имунен отговор. Възпаление. Ранно защитен възпалителен отговор.
Етап имунен отговор. Форми на имунен отговор. Възпаление. Ранно защитен възпалителен отговор.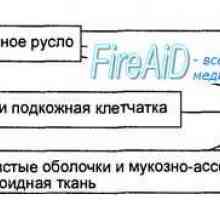 Далак. функция на далака. Лимфни възли. Функциите на лимфните възли.
Далак. функция на далака. Лимфни възли. Функциите на лимфните възли.- Клетъчен протеин TLR5 - ключът за лечение на ревматоиден артрит
 Левкемия, симптоми, лечение, причини
Левкемия, симптоми, лечение, причини